|
|
|
Close Help | ||||||||||||||
|
|
Department of Auditory Neuroscience Head: Prof. MUDr. Josef Syka, DrSc.
Head: Prof. MUDr. Josef Syka, DrSc.Scientists: MUDr. Daniela Buckiova, CSc. Ing. Milan Jilek RNDr. Jiri Popelar, CSc. RNDr. Natalia Rybalko, CSc. Dr. Ing. Daniel Suta RNDr. Rostislav Turecek, Ph.D. Ph.D. Students: Mgr. Bohdana Hruskova MUDr. Ladislav Ouda MUDr. Jana Pelanova Mgr. Johana Trojanova Technical Assistants: Jana Janouskova Jan Kolar Jan Setnicka Eva Velebna Address: Videnska 1083, 142 20 Praha 4 Phone: (+420) 296 442 700 Fax: (+420) 296 442 787 E-mail: syka@biomed.cas.cz The main research aim in the Department of Auditory Neuroscience is to investigate the structure and function of the auditory system in animal models under normal conditions and also under different pathological situations such as noise exposure, the administration of ototoxic drugs and stimulation of the auditory system by electrical current. Selected problems are also investigated in man, either in volunteer subjects or in subjects with auditory system pathologies. The experimental approaches are used to solve practical clinical problems such as the diagnosis and treatment of perception deafness and the development of cochlear neuroprostheses. The methods employed range from anatomical and histochemical techniques (tracing of neural tracts by HRP and neurodegenerative techniques, histochemistry of cytochrome oxidase and NADPH-diaphorase, immunocytochemistry of calbindin, parvalbumin and GAD) to electrophysiological techniques (intracellular recording, patch clamp, extracellular recording of single unit activity, recording of evoked potentials with implanted electrodes, recording of DC potentials), behavioral techniques (operant conditioning procedures for estimating hearing functions in laboratory animals) and special audiological and psychoacoustic methods such as the measurement of otoacoustic emissions, the estimation of difference limens for frequency and intensity, and gap function. In addition, methods of nuclear magnetic resonance such as functional magnetic resonance imaging are used in collaboration with the Department of Nuclear Magnetic Resonance of the Institute of Clinical and Experimental Medicine. Studies in experimental audiology in patients are performed in collaboration with the ENT Clinics of the 1st and 2nd Medical Faculties of Charles University in Prague Current research at the Department of Auditory Neuroscience focuses on the following topics: Progress has been made in describing the descending corticotectal auditory pathway. In the rat, descending fibers are axons of the layer V pyramidal cells, and the corticogeniculate pathway originates in layer VI. The descending fibers originate in all three main auditory areas. NADPH-diaphorase-positive neurons are present in the auditory pathway of the rat in subnuclei, which belong to secondary parts of the auditory pathway, i.e. in the external and dorsal cortices of the inferior colliculus and in the medial and suprageniculate nucleus of the medial geniculate body. In the cortex they are dispersed in all layers with the exception of layers I and III. Bilateral ablation of the auditory cortex in rats results in a temporary decrease of NADPH-diaphorase staining in the inferior colliculus. Complementary to the distribution of NADPH-diaphorase, cytochromoxidase has been found in the primary parts of the auditory pathway both in rats and in guinea pigs. A similar complementary distribution was found for the calcium binding proteins calbindin and parvalbumin. For example, in cochlear nuclei calbindin is present in the dorsal cochlear nucleus whereas parvalbumin-positive cells are located in the anteroventral nucleus. Large changes in the thickness of the auditory and visual cortices were observed in very old rats (more than three years old) accompanied by changes in the NADPH-diaphorase neurons. A decrease in the number of NADPH-diaphorase-positive neurons in the inferior colliculus of the rat was described several days after auditory cortex ablation. At the present time the distribution of glutamic-acid decarboxylase (GAD) in the auditory cortex of rats exposed to noise is a subject of study. 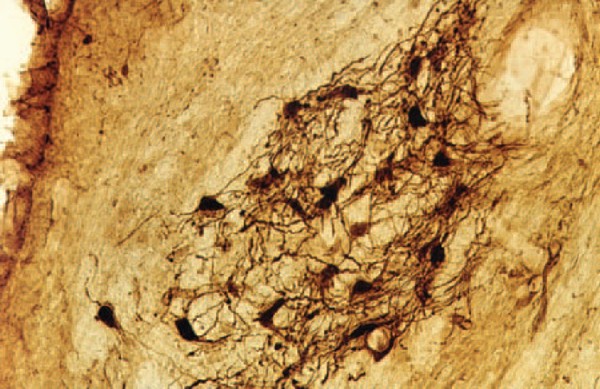 Fig. 1: Calbindin- positive octopus cells in the posteroventral cochlear nucleus of rat Physiology and development of GABA and glycinergic systems in the auditory nuclei The goal of our current projects has been to reveal the mechanisms modulating glutamate release from giant nerve terminals (calyces of Held) in the rat medial nucleus of the trapezoid body (MNTB). Principal cells of the MNTB play a key role in the acoustic information processing by the brain stem auditory circuitry. These cells accurately convert excitatory signals provided by the ventral cochlear nucleus to inhibitory signals directed to the lateral/medial superior olive. Our studies using presynaptic patch-clamp recording provided evidence for the presence of glycine and GABAA receptors in the calyx of Held. These receptors activated by GABA and glycine spillover from the inhibitory fiber endings induced a strongly facilitated glutamate release from the calyx. The mechanism underlying the facilitation appeared to involve a weak depolarization of a nerve terminal, an activation of presynaptic voltage-gated calcium channels, and presynaptic Ca2+ accumulation. We found that in animals younger than postnatal day 11 (P11), and prior to the onset of hearing, the terminals expressed mostly GABAA receptors. After P11, GABAA receptors were largely absent from the terminals and replaced by glycine receptors. 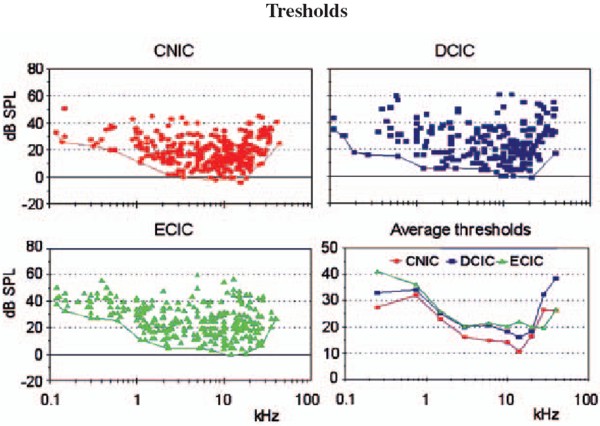 Fig. 2: Mapping of neuronal thresholds to sound stimuli in the inferior colliculus (IC) in guinea pig. 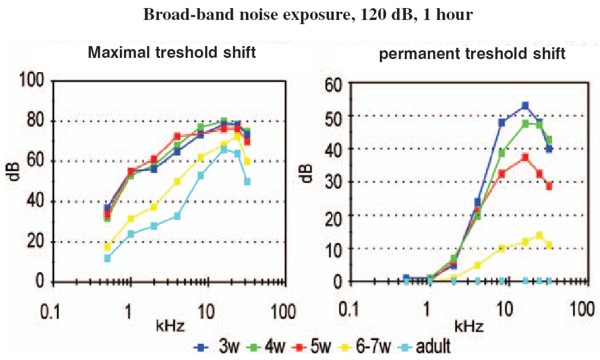 Fig. 3: Threshold shifts after one hour exposure of broad-band noise 120 dB SPL in young and adult rats The role of subcortical auditory nuclei in the processing of acoustical signals Mapping of the parameters of neuronal responses in the inferior colliculus (IC) in guinea pig revealed the tonotopical organisation of this nucleus, which is similar to that in other mammalian species. Isofrequency layers spread through all three subnuclei of the IC; however, the responses of neurons in the "secondary" parts of the IC are different in some parameters from those in the "primary" parts (the sharpness of tuning is smaller, the latencies of responses are longer and the average thresholds are higher). Differences between the secondary and primary parts were also found in the responses of IC neurons to species-specific vocalizations, particularly in cats. However, a high degree of specificity does not exist in the reactions of IC neurons in the guinea pig to vocalizations of its own species. Many of the IC neurons in the guinea pig react to all types of animal vocalizations as well as to noise and pure tones. Similarly, in the medial geniculate body of the guinea pig we did not find specific reactions to species-specific vocalizations, although neurons in the medial geniculate body prefer to respond to the beginning of acoustical stimuli, including vocalizations. Both in the IC and MGB, the spectrotemporal acoustic pattern of vocalization sounds is encoded in a distributed way by a population of neurons. The role of descending pathways in the auditory systemOne of the longstanding topics in the department is the study of the structure and function of the descending pathways in the central auditory system. After performing several morphological studies, a technique of functional ablation of the auditory cortex was introduced to investigate the role of the corticotectal pathway descending from the layer five pyramidal neurons to the inferior colliculi, mostly on the ipsilateral side. The application of the sodium channel blocker tetrodotoxin on the surface of the auditory cortex resulted in the prolongation of the latencies of evoked potentials in the IC, in increases or decreases of the neuronal responses to sound in the IC, as well as in changes in the functional interaction in pairs of IC neurons recorded with one microelectrode. Another study demonstrated that electrical stimulation of distinct loci in the IC produces changes in the distortion product otoacoustic emissions on the ipsilateral side in the guinea pig. Changes in the auditory system in pathological situationsThe exposure of awake, non-anaesthetised guinea pigs to noise results in an enhancement of middle latency responses (MLR) to acoustical stimuli. The amplitude of MLR enhancement is related to the intensity and duration of the noise exposure. The results suggest that the post-exposure amplitude enhancement may be caused by a temporary exhaustion of inhibitory processes in the auditory cortex. Similar effects of noise exposure were observed in rats - a species with typical high frequency hearing. High intensity noise exposure in rats also results in temporary threshold shifts accompanied by enhanced MLR. Recovery functions of both processes (i.e., the threshold shift and the amplitude enhancement) are different, which supports the idea that threshold shifts are generated in the inner ear whereas amplitude enhancement is caused by processes in the auditory cortex or in subcortical nuclei. The effects of noise exposure are even more pronounced in young rats during first five postnatal weeks. Noise exposure, which in adult rats produces a temporary threshold shift, results in young rats (up to the age of 6-7 weeks) in a permanent threshold shift. Histochemical methods also indicate changes in the function of the auditory system after noise exposure. For example, the occurrence of calbindin-positive neurons in dorsal cochlear nuclei increases immediately after noise exposure in rats. Estimation of hearing function in experimental animals with behavioral techniquesOperant conditiong procedures are used to investigate hearing function in rats, with the aim of comparing the results with the results of psychoacoustical tests in man or with the results of audiological tests in patients. Hearing sensitivity in rats was found to range from 0.5 kHz to 64 kHz, with the lowest threshold values present between 10 and 30 kHz. Frequency discrimination in rats was found to be substantially poorer than in man or in the majority of mammalian species so far tested, with Weber ratios (frequency difference limen divided by frequency) between 3.7 and 7.6%. The Weber ratio in man is around 0.3%. Intensity difference limens were frequency independent and amounted to 2.9 ± 0.5 dB in conditions of intensity increases and 6.5 ± 1.6 dB in conditions of intensity decreases. Temporal resolution in rats was measured by detection of gaps in a continuosly presented sound (gap function). Thresholds for gap detection were found to be in the range of 2-3 ms, which is comparable with values found in man. In rats, only very small changes in gap detection threshold were observed after bilateral ablation of the auditory cortex, which contrasts with the widely accepted hypothesis that the auditory cortex plays an important role in gap detection. Studies of hearing function in manIn young volunteers exposed to 85 dB SPL white noise or music of the same intensity, greater activation of the temporal lobes was observed with the musical signal. The laterality of auditory cortex activation by music was significantly different when the volunteers listened to either instrumental music or the same music with the addition of a singer´s voice. Temporary threshold shifts and a significant decline in transient, distortion product and spontaneous otoacoustic emissions were observed in young volunteers exposed for four hours to amplified music (97 dB SPL /A/) previously recorded in a discotheque. The hearing abilities of a group of elderly subjects were compared with a group of young normal hearing volunteers. The results support the view that presbycusis in man is produced by a combination of declining function of the auditory periphery and deteriorated function of the central auditory system. Relevant publications before 1998 1. Johnstone, B.M., Patuzzi, R., Syka, J., Sykova, E. (1989) Stimulus-related potassium changes in the organ of Corti of guinea pig. J. Physiol. (London) 408: 77-92 2. Syka, J. (1989) Experimental models of sensorineural hearing loss. Effects of noise and ototoxic drugs on hearing. Prog. Sensory Physiol. 9: 97-170 3. Schrott, A., Melichar, I., Popelar, J., Syka, J. (1990) Deterioration of hearing function in mice with neural crest defect. Hearing Res. 46: 1-8 4. Taudy, M., Syka, J., Popelar, J, Ulehlova,L. (1992) Carboplatin and cisplatin ototoxicity in guinea pig. Audiology 31, 293-299 5. Ernst, A., Syka, J., Mest, H-J. (1992) Electrophysiological responses of the cochlea to transient asphyxia are influenced by arachidonate metabolites. Prostaglandins 43: 331-338 6. Popelar, J., Syka, J. (1993) Middle latency responses to electrical stimulation of the auditory nerve in unanaesthetised guinea pigs. Hearing Res. 67: 69-74 7. McAnally, K.I., Clark, G.M., Syka, J. (1993) Hair cell mediated responses of the auditory nerve to electrical stimulation of the cochlea in the cat. I. Sinusoidal stimulation. Hearing Res. 67: 55-68 8. Druga, R., Syka, J. (1993) NADPH-diaphorase activity in the central auditory structures of the rat. NeuroReport 4: 999-1002 9. Syka, J., Popelar, J. (1994) Modulation of thresholds to acoustical and electrical stimulation of the intact ear in guinea pig by furosemide and noise. Hearing Res. 75: 1-10 10. Syka, J., Rybalko, N., Popelar, J. (1994) Enhancement of the auditory cortex evoked responses in awake guinea pigs after noise exposure. Hearing Res. 78: 158-168 11. Aitkin, L., Tran, L., Syka, J. (1994) The responses of neurons in subdivisions of the inferior colliculus of cats to tonal, noise and vocal stimuli. Exp. Brain Res. 98: 53-64 12. Popelar, J., Hartmann, R., Syka, J., Klinke, R. (1995) Middle latency responses to acoustical and electrical stimulation of the cochlea in cats. Hearing Res. 92: 63-77 13. Syka, J., Rybalko, N., Brozek, G., Jilek, M. (1996) Auditory frequency and intensity discrimination in pigmented rats. Hearing Res. 100: 107-113 14. Astl, J., Popelar, J., Kvasnak, E., Syka, J. (1996) Comparison of response properties of neurons in the inferior colliculus of guinea pig under different anesthetics. Audiology 35: 335-345 15. Druga, R., Syka, J., Rajkowska, G. (1997) Projections of auditory cortex onto the inferior colliculus in the rat. Physiol. Res. 46: 215-222 Publications 1998-2004 1. Syka, J., Popelar, J., Kvasnak, E., Suta, D. (1998) Processing of vocalization signals in neurons of the infe- rior colliculus and medial geniculate body. In: Central Auditory Processing and Neural Modeling. (Eds. Poon P.W. and Brugge, J.F.) Plenum Press, New York, pp. 1-11 2. Syka, J., Popelar, J., Kvasnak, E., Suta, D. (1998) Processing of simple and complex acoustical signals in the inferior colliculus and medial geniculate body of the guinea pig. In: Psychophysical and Physiological Advances in Hearing. (Eds. A.R. Palmer, A. Rees, A.Q. Summerfield, R. Meddis) Whurr Publishers, London, pp. 491-496 3. Popelar, J., Valvoda, J., Syka, J. (1999) Acoustically and electrically evoked contralateral suppression of otoacoustic emissions in guinea pigs. Hearing Res. 135: 61-70 4. Syka, J., Rybalko, N. (2000) Threshold shifts and enhancement of cortical evoked responses after noise exposure in rats. Hearing Res. 139: 59-68 5. Kvasnak, E., Suta, D., Popelar, J., Syka, J. (2000) Neuronal connections in the medial geniculate body of the guinea pig. Exp. Brain Res. 132: 87-102 6. Kvasnak, E., Popelar, J., Syka, J. (2000) Discharge properties of neurones in subdivisions of the medial geniculate body of the guinea pig. Physiol. Res. 49: 369-378 7. Syka, J., Popelar, J., Kvasnak, E., Astl, J. (2000) Response properties of neurons in the central nucleus and external and dorsal cortices of the inferior colliculus in guinea pig. Exp. Brain Res. 133: 254-266 8. Turecek, R., Trussell, L.O. (2000) Control of synaptic depression by glutamate transporters. J. Neurosci. 20: 2054-2063 9. Turecek, R., Trussell, L.O. (2001) Presynaptic glycine receptors enhance transmitter release at a mam- malian central synapse. Nature 411: 587-590 10. Popelar, J., Erre, J.-P., Syka, J., Aran, J.-M. (2001) Effects of contralateral acoustical stimulation on three measures of cochlear function in the guinea pig. Hearing Res. 152: 128-138 11. Rybalko, N., Syka, J. (2001) Susceptibility to noise exposure during postnatal development in rats. Hearing Res. 155: 32-40 12. Druga, R., Syka, J. (2001) Effect of auditory cortex lesions on NADPH-diaphorase staining in the inferior colliculus of rat. NeuroReport 12: 1555-1559 13. Mazelova, J., Valvoda, J., Popelar, J., Syka, J. (2001) The influence of single exposure to amplified music on hearing of young listeners. In: Noise Induced Hearing Loss: Basic Mechanisms, Prevention and Control. D.Henderson, D.Prasher, R.Kopke, R.Salvi and R.Hamernik (Eds.). NRN Publications, London, pp. 365-375 14. Syka, J. (2002) Plastic changes in the central auditory system after hearing loss, restoration of function and learning. Physiol. Rev. 82: 601-636 15. Turecek, R., Trussell, L.O. (2002) Reciprocal developmental regulation of presynaptic ionotropic receptors. Proc. Natl. Acad. Sci. USA 99: 13884-13889 16. Popelar, J., Mazelova, J., Syka, J. (2002) Effects of electrical stimulation of the inferior colliculus on 2f1-f2 distortion product otoacoustic emissions in anesthetized guinea pigs. Hearing Res. 170: 116-126 17. Syka, J., Rybalko, N., Mazelova, J., Druga, R. (2002) Gap detection threshold in the rat before and after auditory cortex ablation. Hearing Res. 172: 151-159 18. Nwabueze-Ogbo, F.C., Popelar, J., Syka, J. (2002) Changes in the acoustically evoked activity in the inferi- or colliculus of the rat after functional ablation of the auditory cortex. Physiol. Res. 51: S95-S104 19. Mazelova, J., Popelar, J., Syka, J. (2003) Auditory function in presbycusis: peripheral vs. central changes. Exp. Gerontol. 38: 87-94 20. Popelar, J., Nwabueze-Ogbo, F.C., Syka, J. (2003) Changes in neuronal activity of the inferior colliculus in rat after temporal inactivation of the auditory cortex. Physiol. Res. 52: 615-628 21. Ouda, L., Nwabueze-Ogbo, F.C., Druga, R., Syka, J. (2003) NADPH-diaphorase - positive neurons in the auditory cortex of young and old rats. NeuroReport 14: 363-366 22. Suta, D., Kvasnak, E., Popelar, J., Syka, J. (2003) Representation of species-specific vocalizations in the inferior colliculus of the guinea pig. J. Neurophysiol. 90: 3794-3808 23. Popelar, J., Groh, D., Mazelova, J., Syka, J. (2003) Cochlear function in young and adult Fischer F344 rats. Hear. Res. 186: 75-84 24. Glogarova, K., Buckiova, D. (2004) Changes in sialylation in homozygous Sp2H mouse mutant embryos. Birth Defects Res Part A Clin Mol Teratol. 70: 142-52 25. Kyselova, V., Peknicova, J., Boubelik, M., Buckiova, D. (2004) Body and organ weight, sperm acrosomal status and reproduction after genistein and diethylstilbestrol treatment of CD1 mice in a multigenerational study. Theriogenology 61: 1307-1325 26. Awatramani, G.B., Turecek, R., Trussell, L.O. (2004) Inhibitory control at a synaptic relay. J Neurosci. 24: 2643-2647 27. Turecek, R., Vlcek, K., Petrovic, M., Horak, M., Vlachova, V., Vyklický, L.Jr. (in press) Intracellular spermine decreases open probability of N-methyl-D-aspartate receptor channels. Neuroscience. |